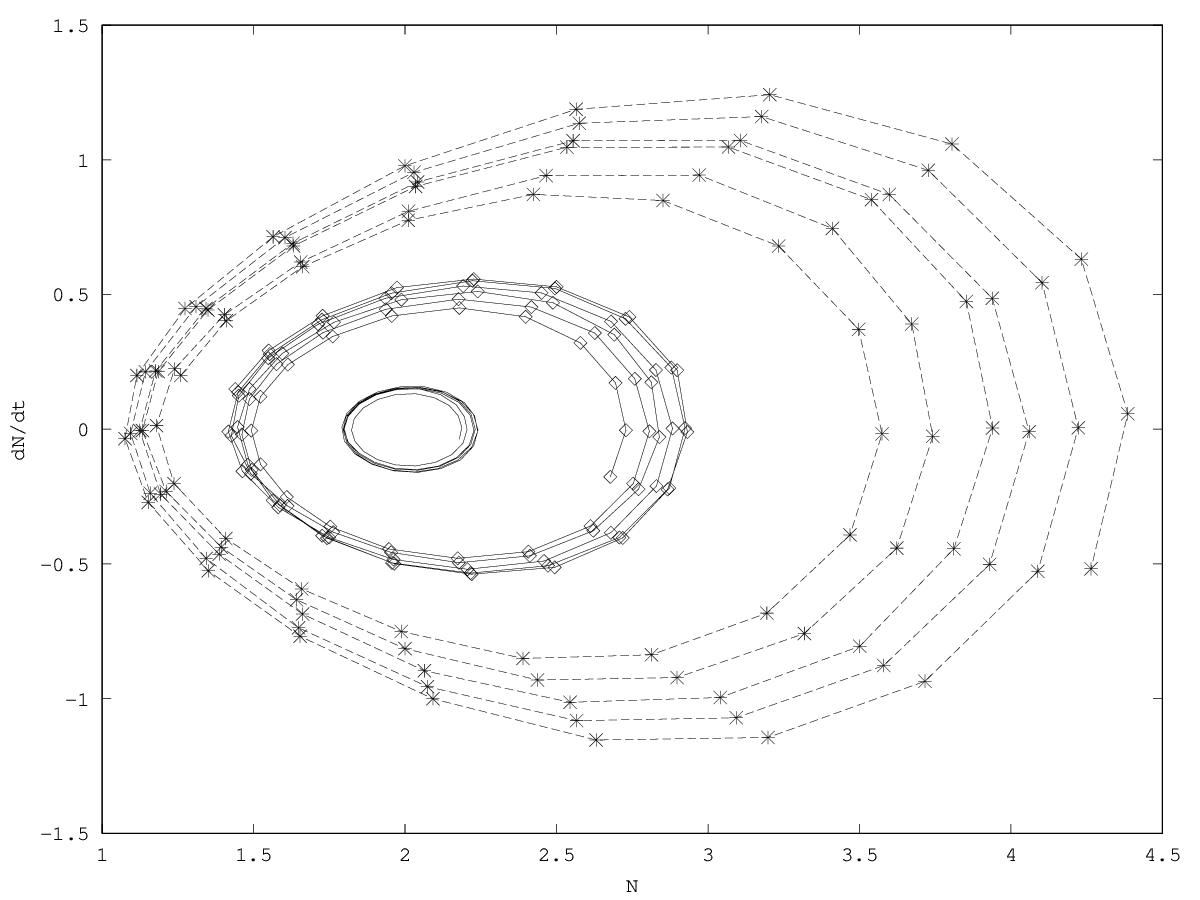
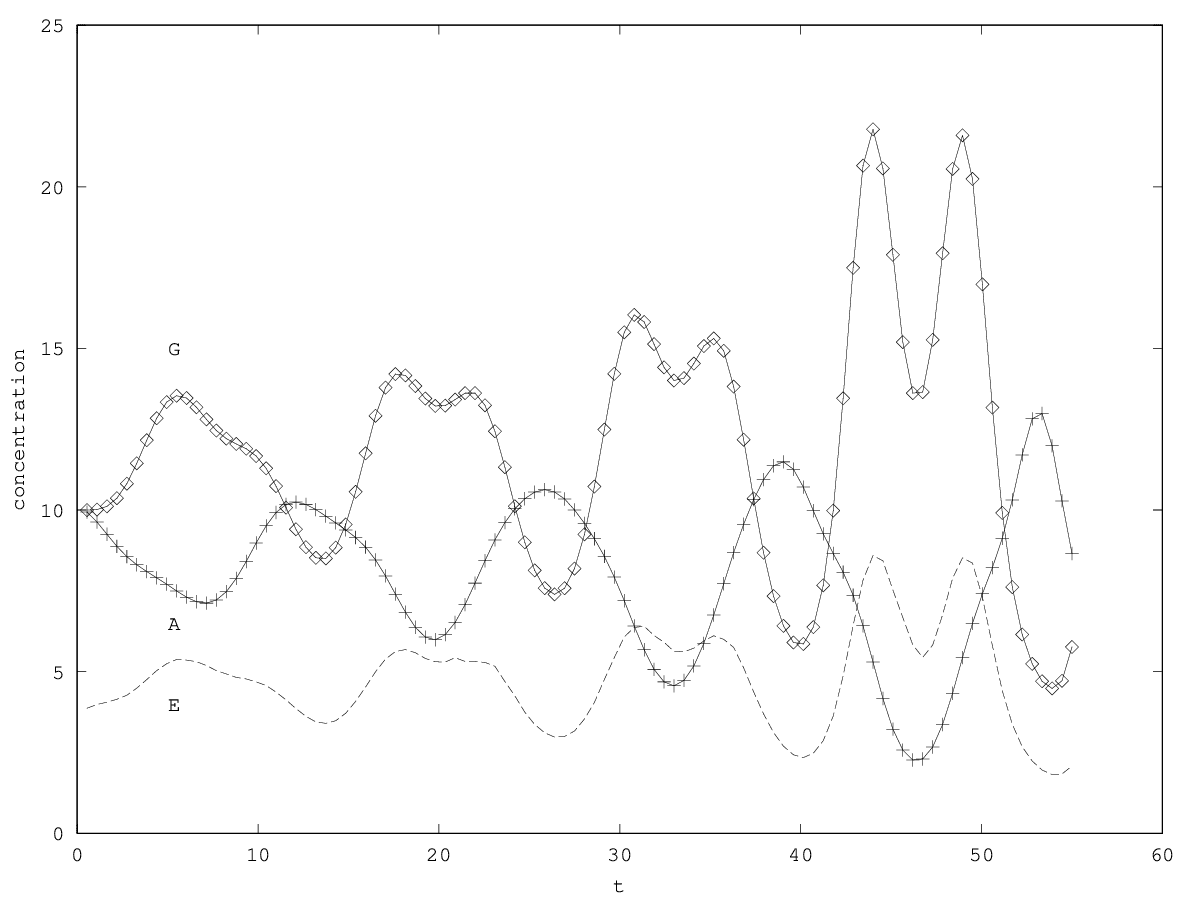
We discuss the perspective of intra-organismal ecology by investigating a family of models of niche construction. We consider first and second order models.
Abstract
In this article, we discuss the perspective of intraorganismal ecology by investigating a family of ecological models. We consider two types of models. First-order models describe the population dynamics as being directly affected by ecological factors (here understood as nutrients, space, etc). They might be thought of as analogous to Aristotelian physics. Second-order models describe the population dynamics as being indirectly affected, the ecological factors now affecting the derivative of the growth rate (that is, the population acceleration), possibly through an impact on nongenetically inherited factors. Second-order models might be thought of as analogous to Galilean physics. In a companion article, we apply these ideas to a situation of gene therapy.
We apply the perspective of intra-organismal ecology by investigating a family of ecological models suitable to describe a gene therapy.
Abstract
In this article, we apply the perspective of intraorganismal ecology by investigating a family of ecological models suitable to describe a gene therapy for a particular metabolic disorder, the adenosine deaminase deficiency. The gene therapy is modeled as the prospective ecological invasion of an organ (here, bone marrow) by genetically modified stem cells, which then operate niche construction in the cellular environment by releasing an enzyme they synthesize. We show that depending on the order chosen for the model (a choice that cannot be made on a priori assumptions), different kinds of dynamics are expected, possibly leading to different therapeutic strategies. This drives us to discuss several features of the extension of ecology to intraorganismal ecology.
This note introduces work in Theoretical Biology in the book: Perspectives on Organisms: Biological Time, Symmetries and Singularities.
Abstract
This note introduces recent work in Theoretical Biology by borrowing from the Introduction (chapter 1) of the book by the authors: "Perspectives on Organisms: Biological Time, Symmetries and Singularities", Springer, 2014. The idea is to work towards a Theory of Organisms analogue and along the Theory of Evolution, where ontogenesis could be considered as part of phylogenesis. As a matter of fact, the latter is made out of "segments" of the first: phylogenesis is the "sum" of ontogenetic paths and they should be made intelligible by similar principles. To this aim, we look at ontogenesis from different perspectives. By this, we shed light on the unity of the organism from different points of view, yet constantly keeping that unity as a core invariant. The analysis of invariance, as the result of theoretical symmetries, and of symmetry changes, is a key theme of the approach in the book and in the discussion in this note.
In 1999, C Sonnenschein and AM Soto proposed the tissue organization field theory (TOFT). The TOFT posits 1) that cancer is a tissue-based disease whereby carcinogens (directly) and germ-line mutations (indirectly) alter normal interactions between the stroma and adjacent epithelium; and 2) the default state of all cells is proliferation with variation and motility. This later premise is relevant to and compatible with evolutionary theory, and is diametrically opposed to that of the somatic mutation theory. This theoretical change is incompatible with the reductionist genocentric perspective generated by the molecular biology revolution. Rather than forcing a “bricolage” we decided to frontally attack the problem by joining efforts with philosophers, mathematicians and theoretical biologists to search for principles upon which to build a theory of organisms (this work is currently supported by the Pascal Chair). While the theory of evolution has provided an increasingly adequate explanation of phylogeny, biology still lacks a theory of organisms that would encompass ontogeny and life cycles, and thus phenomena on a conception to death time-scale. To achieve this goal we propose that theoretical extensions of physics are required in order to grasp the living state of matter. Such extensions will help to describe the proper biological observables, i.e. the phenotypes. Biological entities must also follow the underlying principles used to understand inert matter. However, these physical laws and principles may not suffice to make the biological dynamics intelligible at the phenotypic level. By analogy with classical mechanics, where the principle of inertiais the default state of inert matter, we are proposing two aspects of the default state in biology, and a framing principle, namely: i) Default state: cell proliferation with variation as a constitutive property of the living. Variation is generated in particular by the mere fact that cell division generates two overall similar, but not identical cells. ii) Default state: motility, which encompasses cell and organismic movements as well as movement within cells. iii) Framing principle: life phenomena exhibit never identical iterations of a morphogenetic process. Organisms are the consequence of the inherent variability generated by proliferation, motility and auto-organization which operate within the framing principle. From these basic premises, we will elaborate on the generation of robustness, the structure of theoretical determination, and the identification of biological proper observables.
Universidad del Pais Vasco (EHU/UPV) Campus de Guipuzkoa
The workshop will discuss the notion of biological organization from a systemic- perspective. In particular it will focus on its intrinsic hierarchical dimension, and on the role organization plays in the understanding of the transition from pre-biotic to minimal living systems and of more complex forms of biological, cognitive and ecological systems.
• Lower type 1 collagen concentra1on (0.5 mg/ml) increases elonga1on of structures compared to 1 mg/ml. • Fibroblasts enhances elonga1on of structures in 1 mg/ml, but not 0.5 mg/ml. • In 1 mg/ml collagen, elongated epithelial structures had lumen forma1on when co-‐cultured with fibroblasts. • Hormones in co-‐cultures of T47D + fibroblasts, significantly altered the phenotype in terms of elonga1on.
In vitro 3D simulations of biological systems are critical for understanding morphogenesis and patterns of normal and abnormal development in tissues. Here we present SAMA, a novel method through which epithelial structures grown in 3D cultures can be imaged, reconstructed in 3D and analyzed with minimum human intervention and, therefore, bias. SAMA gives us an accurate picture of the epithelial structures and hence, more information compared to 2D morphometric analysis. In addition, this automated method is less time consuming than regular 2D morphometrics.
The coupling of generic objects, defined by their symmetries, on the one side, and the corresponding trajectories, on the other side, is at the core of the objectivation of phenomena understood in existing physical theories. As for proper biological phenomena, we hypothesize that they do not possess stable theoretical symmetries. Thus, biological objects are specific in the sense that they are the result of historical and contingent changes of symmetries. This provides a specific notion of randomness, proper to biological phenomena.
In this talk, we will discuss some propositions of extensions of physical theories to understand biological objects. we will focus on some aspects of biological time. First, we will focus on biological rhythms, in relationship with allometric relations. Then we will discuss the notions of biological complexity and organization, in the context of macroevolution.
Bulk properties do not determine shape; however, localized regions of collagen fiber alignment are required for ductal elongation and branching.
Abstract
Background: Mammary gland morphogenesis involves ductal elongation, branching, and budding. All of these processes are mediated by stroma - epithelium interactions. Biomechanical factors, such as matrix stiffness, have been established as important factors in these interactions. For example, epithelial cells fail to form normal acinar structures in vitro in 3D gels that exceed the stiffness of a normal mammary gland. Additionally, heterogeneity in the spatial distribution of acini and ducts within individual collagen gels suggests that local organization of the matrix may guide morphogenesis. Here, we quantified the effects of both bulk material stiffness and local collagen fiber arrangement on epithelial morphogenesis. Results: The formation of ducts and acini from single cells and the reorganization of the collagen fiber network were quantified using time-lapse confocal microscopy. MCF10A cells organized the surrounding collagen fibers during the first twelve hours after seeding. Collagen fiber density and alignment relative to the epithelial surface significantly increased within the first twelve hours and were a major influence in the shaping of the mammary epithelium. The addition of Matrigel to the collagen fiber network impaired cell-mediated reorganization of the matrix and increased the probability of spheroidal acini rather than branching ducts. The mechanical anisotropy created by regions of highly aligned collagen fibers facilitated elongation and branching, which was significantly correlated with fiber organization. In contrast, changes in bulk stiffness were not a strong predictor of this epithelial morphology. Conclusions: Localized regions of collagen fiber alignment are required for ductal elongation and branching suggesting the importance of local mechanical anisotropy in mammary epithelial morphogenesis. Similar principles may govern the morphology of branching and budding in other tissues and organs.
Keywords: Collagens, Morphogenesis, Extracellular matrix, Gels, Anisotropy, Stiffness, Scanning electron microscopy, Mammary gland development
The core of the objectivization in physics is the coupling of generic objects, as defined by their symmetries, and their corresponding specific trajectories. As for biological phenomena, we posit that they do not possess stable theoretical symmetries. Thus, biological objects are specific in the sense that they are the result of historical and contingent changes of symmetries. This situation will allow us to discuss a specific notion of measurement for biological objects: measurements in biology co-establishes and often stabilize the theoretical symmetries of biological objects.
This authored monograph introduces a genuinely theoretical approach to biology. Starting point is the investigation of empirical biological scaling including their variability, which is found in the literature, e.g. allometric relationships, fractals, etc. The book then analyzes two different aspects of biological time: first, a supplementary temporal dimension to accommodate proper biological rhythms; secondly, the concepts of protension and retention as a means of local organization of time in living organisms. Moreover, the book investigates the role of symmetry in biology, in view of its ubiquitous importance in physics. In relation with the notion of extended critical transitions, the book proposes that organisms and their evolution can be characterized by continued symmetry changes, which accounts for the irreducibility of their historicity and variability. The authors also introduce the concept of anti-entropy as a measure for the potential of variability, being equally understood as alterations in symmetry. By this, the book provides a mathematical account of Gould’s analysis of phenotypic complexity with respect to biological evolution. The target audience primarily comprises researchers interested in new theoretical approaches to biology, from physical, biological or philosophical backgrounds, but the book may also be beneficial for graduate students who want to enter this field.
Abstract
This authored monograph introduces a genuinely theoretical approach to biology. Starting point is the investigation of empirical biological scaling including their variability, which is found in the literature, e.g. allometric relationships, fractals, etc. The book then analyzes two different aspects of biological time: first, a supplementary temporal dimension to accommodate proper biological rhythms; secondly, the concepts of protension and retention as a means of local organization of time in living organisms. Moreover, the book investigates the role of symmetry in biology, in view of its ubiquitous importance in physics. In relation with the notion of extended critical transitions, the book proposes that organisms and their evolution can be characterized by continued symmetry changes, which accounts for the irreducibility of their historicity and variability. The authors also introduce the concept of anti-entropy as a measure for the potential of variability, being equally understood as alterations in symmetry. By this, the book provides a mathematical account of Gould’s analysis of phenotypic complexity with respect to biological evolution. The target audience primarily comprises researchers interested in new theoretical approaches to biology, from physical, biological or philosophical backgrounds, but the book may also be beneficial for graduate students who want to enter this field.